
Внутренний скелет Aranei, как у других паукообразных и Xiphosura, состоит из образований двоякого происхождения, С одной стороны, имеются аподемы, или эндосклериты (эндапофизы), т. е. более или менее глубокие впячивания наружной хитиновой кутикулы, с другой — эндохондриты, или скелетные образования мезодермальной природы (Schimkewitsch, 1893; Pereyaslawzewa, 1907), часто не связанные с наружным хитином.
Аподемы
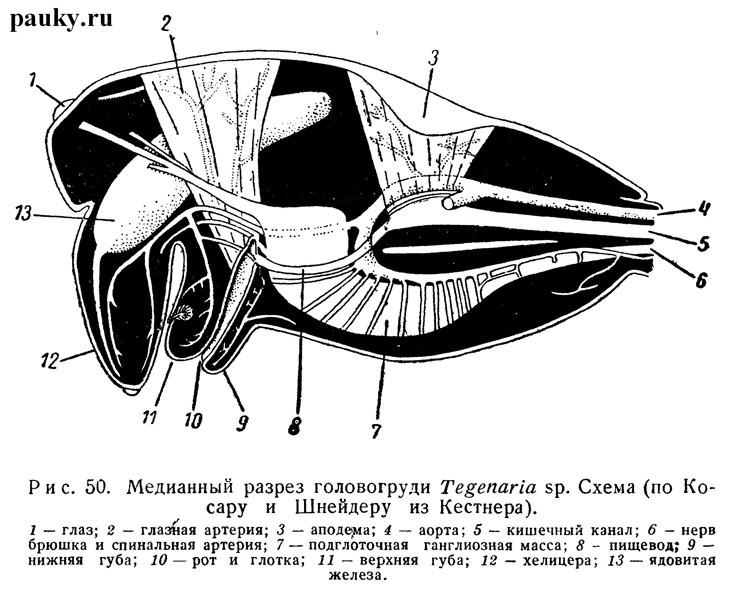
Аподемы головогруди развиты очень слабо. Небольшую непарную аподему образует карапакс в области срединной ямки (см. рис. 50); к ней прикрепляются мускулы сосательного желудка. На брюшной стороне головогруди небольшие пластинчатые аподемы обыкновенно развиты лишь на тазиках педипальп.
В стебельке задние углы тергальной пластинки — лорума снабжены маленькими аподемальными отростками. Ниже их, на границе стебелька и брюшка, лежат очень маленькие аподемы, к которым прикрепляются медиальные продольные мускулы брюшка. Но особенный интерес представляют вентральные аподемы брюшка, так как они развиваются на задних границах сегментов и за счет некоторых из них формируется часть трахей. Это довольно значительные языкообразные или пластинчатые впячивания интерсегментарных участков кутикулы (рис.43).
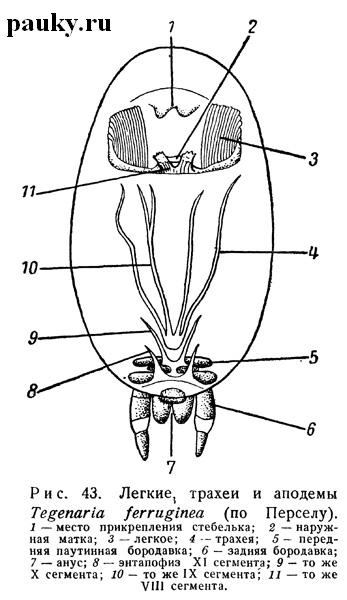
Передние аподемы помещаются на уровне легочных стигм (у четырехлегочных пауков — на уровне стигм первой пары легких) и представляют собой придатки эпигастрального впячивания, принадлежащего VIII сегменту. Следующая пара аподем IX сегмента лежит позади трахей (или второй пары легких) и, наконец, две пары аподемальных впячиваний, относящихся к X и XI сегментам, помещаются позади передних и задних паутинных бородавок.
Эндохондриты
Эндохондриты представлены спинными элементами — эндотергитами (endoterga) и брюшными — эндостернитами (еndosterna).
Единственный и вместе с тем самый крупный из эндохондритов — непарный эндостернит головогруди, отвечающий таковому Limulus и других Arachnida, лежит между сосательным желудком и подглоточной нервной массой. Вероятно, он образовался в результате слияния нескольких метамерных эндостернитов, сходно со стернальной пластинкой, Возникшей путем слияния нескольких стернитов. Эта довольно широкая, большая бесцветная пластинка хрящеватой консистенции обычно вогнута сверху и спереди со стороны головного мозга глубоко вырезана (рис.44).
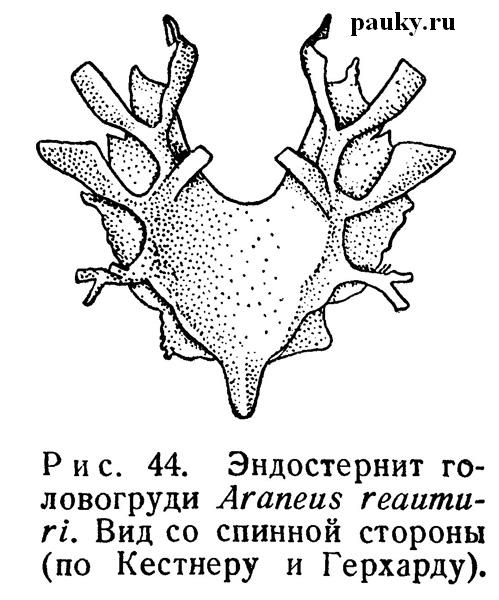
На ее поверхности развиваются пластинчатые выросты. К ним прикрепляются мощные мускулы, связывающие эндостернит со спинным щитом. От брюшной поверхности эндостернита отходят мышцы к стернальной пластинке. Спинная поверхность эндостернита связана мускульными пучками с сосательным желудком.
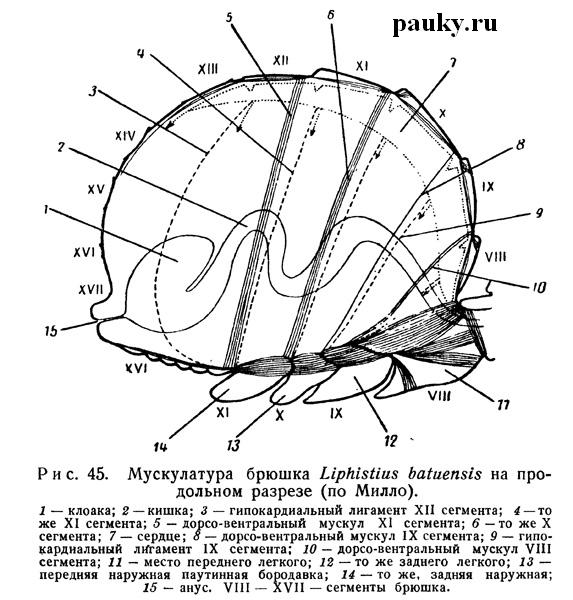
В брюшке эндохондриты представлены парными элементами, сохраняющими сегментарное расположение. Наиболее полно они развиты у Liphistiidae, у которых, дорсально между каждыми двумя тергальными пластинками имеется пара небольших эндотергитов (рис. 45).
На брюшной стороне им отвечают парные медиальные эндостерниты, связанные попарно поперечными сухожильными тяжами и прикрепляющиеся к аподемам своего сегмента. Кроме того, в каждом сегменте имеется еще пара латеральных эндостернитов (рис. 46). К эндохондритам прикрепляется мускулатура.
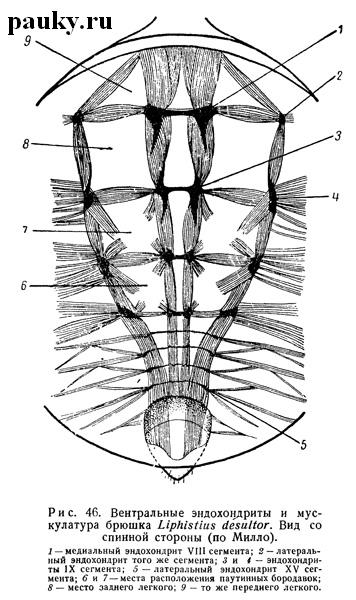
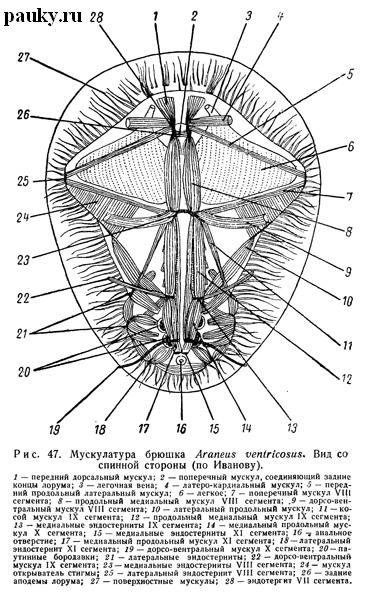
Все эндохондриты брюшка Liphistiidae имеют строго метамерное расположение, помещаясь на задних границах своих сегментов. Однако в сегментах, лежащих позади задних паутинных бородавок и подвергшихся уже значительной редукции, они малы и недоразвиты, что особенно хорошо видно на эндостернитах. У всех прочих пауков эти задние эндостерниты не развиваются вовсе (рис. 47), что связано с полной редукцией вентральных отделов соответствующих сегментов. Эндотергиты также исчезают полностью или частично. В последнем случае они сильно смещены.
Мускулатура пауков
Мускулатура пауков известна по старым работам (Kessler, 1849; Blanchard, 1860; Schimkewitsch, 1884). Она особенно высоко дифференцирована в головогруди. Здесь, помимо мышц конечностей, заслуживают упоминания мускулы, связывающие карапакс и стернум с эндостенитом, мускулы сосательного желудка, глотки и верхней губы (рис. 50).
Мускулы конечностей, недавно очень подробно изученные Германом (Hermann, 1957) у Zygiella x-notata, лежат частью в головогруди, частью в члениках самих конечностей.
Первые осуществляют движение тазиков; они прикрепляются или к эндостерниту, или к боковым стенкам головогруди. Наиболее дифференцирована мускулатура хелицер; каждая хелицера снабжена 21 мышцей, из коих большая часть прикрепляется к хитину передней части головогруди. Кроме того, в основном членике хелицеры помещаются два сгибателя и два разгибателя концевого членика.
Несколько мускульных пучков соединяют головогрудь с брюшком. В стебельке, рядом с кишечником, проходит пара продольных мускулов, которые спереди начинаются в головогруди от эндостернита, а сзади прикрепляются к спинным мускулам первого сегмента брюшка. Более тонкие парные мускульные тяжи тянутся на вентральной стороне стебелька, прикрепляясь сзади к аподемам VII сегмента. Кроме того, в стебельке имеются две пары дорсо-вентральных мышц.
В брюшке, непосредственно под мягкими покровами, прикрепляясь к ним во многих местах, лежит сетчатый слой мускульных волокон (рис. 47). Будучи развит весьма неодинаково в разных семействах, он обычно образует рыхлый мускульный мешок, окружающий внутренности (аналогично кожно-мускульному мешку червей). Его сокращения, вероятно, имеют важное значение для циркуляции крови в брюшке и способствуют выдавливанию пищевых масс из долек печени. От висцеральной массы он отделен щелевидным просветом кровеносного синуса.
Но совершенно особый интерес представляет сохраняющая еще метамерный характер глубже лежащая мускулатура брюшка, строение которой чрезвычайно важно для понимания архитектоники этого отдела тела. Наиболее примитивное состояние брюшной мускулатуры свойственно Liphistiidae с их хорошо расчлененным брюшком. У этих пауков эндотергиты и медиальные эндостерниты четырех передних сегментов брюшка (VIII—XI туловищных сегментов) соединены дорсо-вентральными мускулами (рис. 45), располагающимися, следовательно, интерсегментарно.
Эндотергиты связаны друг с другом слабыми парными продольными дорсальными мышцами, которые в задних сегментах сближаются. На брюшной стороне продольные вентральные мышцы (рис. 46) соответственным образом соединяют эндостерниты соседних сегментов, образуя четыре продольных ряда: два медиальных и два латеральных. Кроме того, эндостерниты каждого сегмента соединяются вентральными поперечными мускульными пучками, за исключением медиальных эндостернитов четырех передних сегментов, которые связаны сухожильными тяжами.
У всех остальных пауков мускулатура брюшка значительно редуцируется, сохраняя, однако, такой же метамерный характер.
- Во-первых, в связи с редукцией задних сегментов сохраняются только мышцы, принадлежащие четырем передним сегментам.
- Во-вторых, в громадном большинстве случаев имеет место дальнейшая редукция и в этих сегментах; она касается различных мышц и выражена в различной степени в разных семействах.
- В-третьих, сохраняющиеся мышцы испытывают значительные смещения, утрачивая свое первоначальное расположение. Все эти изменения связаны с возрастающей интеграцией брюшка.
По степени редукции брюшной мускулатуры все Mygalo- и Araneomorphae могут быть разделены на несколько групп (Millot, 1936а; Rasmont, 1952). Очень примитивные, близкие к Liphistiidae отношения свойственны Eresidae и Uloboridae. Так, у Eresus из вентрального комплекса мышц четырех сегментов отсутствуют только поперечные мускулы между медиальными эндостернитами и сохраняются все четыре пары дорсо-вентральных мускулов. У других форм редуцируются отдельные группы продольных мышечных пучков и в различной степени — дорсо-вентральные мускулы. Последние развиты в числе трех пар (Araneidae —рис. 47, Tetragnathidae, Hypochilidae, Urocteidae и др.), двух или одной пары (Theridiidae, Thomisidae, Lycosidae, Salticidae и др.) или даже вовсе отсутствуют (Dysderidae, Sicariidae, Pholcidae).
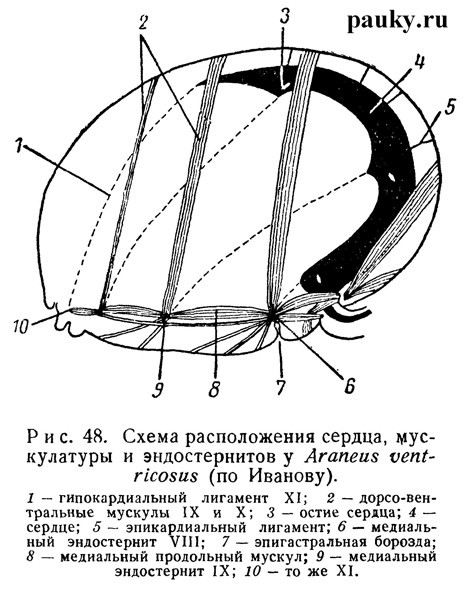
Очень характерны для Araneomorphae вторичные смещения мышц. Так, очень сильно варьирует положение мускулов, связанных с медиальными эндостернитами обычно сильно удлиненного IX (второго брюшного) сегмента. Эти эндостерниты лежат на языкообразных аподемах и в зависимости от длины и положения последних сильно смещены у разных родов то вперед, то назад. У Liphistiidae дорсо-вентральные мускулы параллельны друг другу и занимают интерсегментарное положение, а верхние концы их лежат в одной плоскости с остиями сердца. Но у Araneomorphae спинные концы этих мышц вторично смещаются, а сердце укорачивается, и в результате первичные отношения мышц и остий нарушаются (рис. 48).
По данным Кроме (1955), паукам, подобно большинству паукообразных, свойственна сегментированная продольная дорсальная мускулатура брюшка, несмотря на то, что внешне этот отдел тела не расчленен. Первоначально каждый сегмент брюшка имел пару коротких продольных спинных мускулов. Эти отношения сохранились только у Liphistius и Hypochilus, у остальных пауков VIII и IX сегменты не имеют уже типичных метамерных мускулов, которые большей частью, видоизменяются в так называемые латеральные кардиальные мускулы, проникающие в область стебелька.
Подвижность паутинных бородавок обеспечивается мускульными пучками (рис. 47), прикрепляющимися к базальному членику и связанными с эндохондритами X и XI сегментов. Кроме того, в самих бородавках имеются мускулы, осуществляющие движение их второго членика (в средних бородавках они отсутствуют). Анальный бугорок имеет пару собственных маленьких мускулов.





