
Яйца пауков очень богаты желтком и относятся к центролецитальному типу. Помимо первичной, или желточной, оболочки, их одевает хорион, выделяемый в половых путях сайки. Яйца относительно велики, особенно у крупных видов (например, у тарантула — Lycosa singoriensis — 2 мм в диаметре).
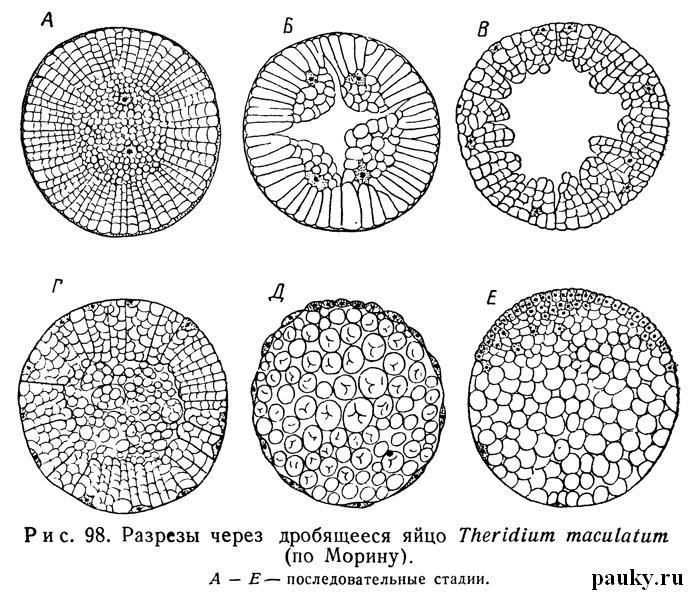
Ядро яйца занимает центральное положение среди многочисленных желточных зерен и окружено участком плазмы, от которого к периферии тянутся плазматические тяжи, сливающиеся под желточной оболочкой с тонким поверхностным слоем бластемы (рис. 98,А). В свежеотложенных яйцах полярность не выражена, но уже через 1—2 часа желток, просто в силу большего удельного его веса, начинает концентрироваться в нижней половине яйца (Заленский, Бальбиани, Лоси).
Дробление
Дробление, как у большинства паукообразных, поверхностное. Сперва ядро делится последовательно на 2, 4, 8 и 16 дочерних ядер, окруженных каждое своим участком плазмы. Уже при первых делениях ядра желточные зерна ориентируются радиально или по крайней мере обнаруживают тенденцию к такому расположению (рис. 98).
С образованием двух ядер вся желточная масса распадается на две половины и нередко (например, у Lycosa и Segestria) настолько полно, что создается ложное впечатление тотального разделения яйца на два бластомера.
Дальнейшие деления ядер также сопровождаются распадением желточной массы и группированием около каждого ядра своего участка желтка — так называемой розетки (Philodromus, Agelena, Segestria). На стадии 16—32—64 ядер последние выходят на поверхность яйца. Розетки образуют теперь так называемые желточные пирамиды (рис. 98,Б). Одновременно в середине яйца появляется обширная полость, наполненная жидкостью и не совсем правильно называемая полостью дробления, или бластоцелем.
Хотя яйцо на этой стадии и напоминает настоящую бластулу, возникшую в результате тотального дробления, но клеточных границ между желточными пирамидами никогда не бывает.
Объединение желточных зерен в одну пирамиду объясняется, по-видимому, лишь силами сцепления, свойственными примыкающей к ядру плазме. Поэтому, вопреки некоторым авторам (Korschelt, Heider, 1892; Давыдов, 1914, Dawydoff, 1949), о тотальном дроблении у пауков не может быть речи.
На стадии около 100 клеток желточные пирамиды сливаются, желток отделяется от плазматического слоя бластодермы, и происходит замечательное явление сжатия зародыша, описанное впервые Заленским (1871). При этом бластулообразный зародыш уменьшается примерно на одну треть своего первоначального объема. В то же время между бластодермой и яйцевой оболочкой накапливается прозрачная перивителлиновая жидкость, выступающая из яйца, а полость бластоцеля заполняется массой желтка (Holm, 1940).
Сжатие происходит в течение 5 — 6 часов и приводит к весьма важным результатам. Вокруг зародыша, несмотря на отсутствие зародышевых оболочек, создается пространство, необходимое для свободного его развития. С другой стороны, это пространство, наполненное жидкостью, играет ту же роль, что и амниотическая полость с ее жидкостью у насекомых и наземных позвоночных, т. е. предохраняет зародыш от высыхания и вообще имеет защитное значение. Таким образом, сжатие зародыша у пауков — этих типично наземных членистоногих — есть, прежде всего, приспособление развивающегося яйца к условиям воздушной среды.
Образование первичной пластинки
Итак, выходящие на поверхность яйца ядра образуют бластодерму (см. рис. 98, Д; 99, А). Но часть ядер с участками плазмы остается в периферических слоях желтка в виде разрозненных клеточных элементов — вителлофагов (рис. 98, Е). Сперва бластодерма равномерно одевает все яйцо, но вскоре на одной стороне образуется обширное сгущение клеток бластодермы в виде расплывчатого беловатого пятна — зародышевый диск.
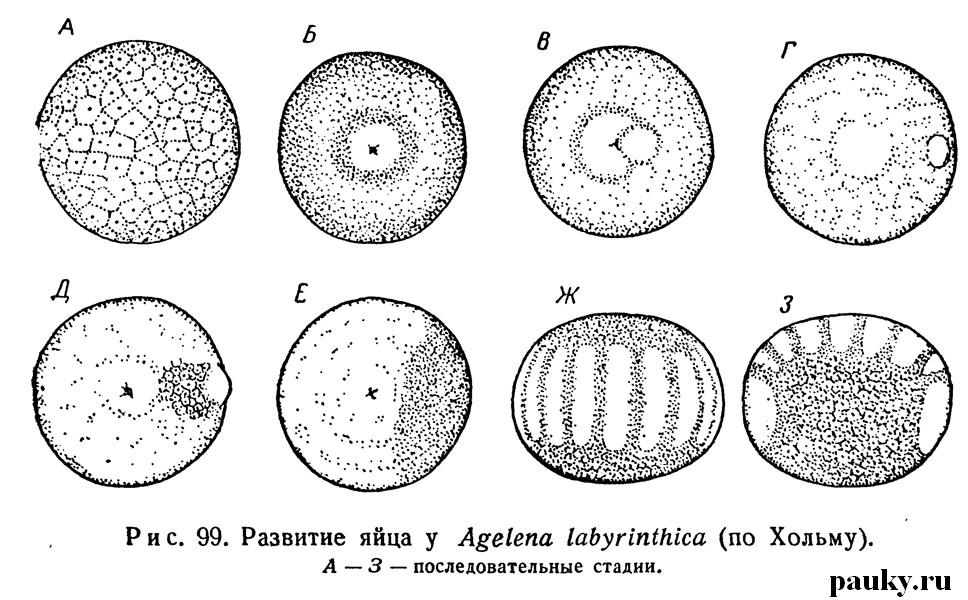
В центре диска появляется маленькая ямка — первичная бороздка, или тастральное впячивание (рис. 99, Д), обнаруженное В. В. Заленским (1871), описанное В. М. Шимкевичем (1887) и изученное недавно Хольмом (Holm, 1940, 1952). Оно отвечает вегетативному полюсу яйца и будущему заднему концу тела (точнее — области презумптивного шестого сегмента брюшка паука).
Из дна бороздки начинают выклиниваться многочисленные клетки, образующие первичную энтодерму. Предполагалось, (Кишинуэ, Гамбургер, Шимкевич и др.), что здесь происходит усиленное размножение клеток. Однако опыты Хольма (Holm, 1952) с маркировкой зародышей Agelena витальными красками показали, что клеточный материал бластодермы перемещается к вегетативному полюсу, и клетки первоначального зародышевого диска постепенно уходят внутрь и дают внутренний листок.
Первичная бороздка является лишь главным очагом выселения клеток, оно начинается еще при образовании самого зародышевого диска и происходит из большей его части. Поэтому очень рано образуется слой внутренних клеток. Двухслойная часть диска называется первичной пластинкой.
Кумулюс
Далее у паука на заднем конце этой пластинки формируется небольшое сгущение энтодермальных клеток, приподнимающее в виде округлого бугорка бластодерму, — кумулюс (cumulus primitivus — рис. 99, В). Довольно скоро кумулюс начинает смещаться на спинную сторону яйца. Сначала он связан еще некоторое время с первичной пластинкой узкой перемычкой, потом между ними образуется все увеличивающееся утонченное поле, состоящее из одного слоя больших уплощенных клеток и получившее название дорсального поля (рис.99, Д, Е).
Зародышевая полоска
Первичная пластинка у пауков затем превращается в зародышевую полоску. Кумулюс, в конце концов, оказывается в передней части зародыша, где вскоре исчезает. Старое мнение о том, что он является зачатком брюшка (Бальфур, Морин, Монтгомери и др.) не подтвердилось. Маркировкой с последующей киносъемкой было показано, что энтодермальные клетки кумулюса перемещаются под слоем бластодермы и частично рассеиваются в области дорсального поля.
Одновременно клетки бластодермы стягиваются на брюшную сторону, где позднее дают эктодерму зародышевой полоски; при этом обнажается слой энтодермы, одевающий желток в области дорсального поля. Поэтому Хольм (Holm, 1952) называет дорсальное поле энтодермальным желточным мешком.
Судьба остальных энтодермальных клеток кумулюса, однако, иная. Позднее, уже на стадии сегментации зародышевой полоски они погружаются внутрь, мигрируют под зачаток брюшка и здесь потом формируют среднюю кишку.
Итак, методом маркировки удалось выяснить, что зародышевый диск является по существу областью бластопора.
Сюда стягиваются клетки бластодермы, и здесь происходит их погружение внутрь и формирование внутренних листков. Когда появляется зародышевый диск, все его клетки представляют будущую энтодерму; вне диска в это время находятся клетки презумптивной энтомезодермы. Погрузившись, энтодерма частью попадает в кумулюс, а частью остается в области первичной полоски, где позднее дает среднюю кишку просомы.
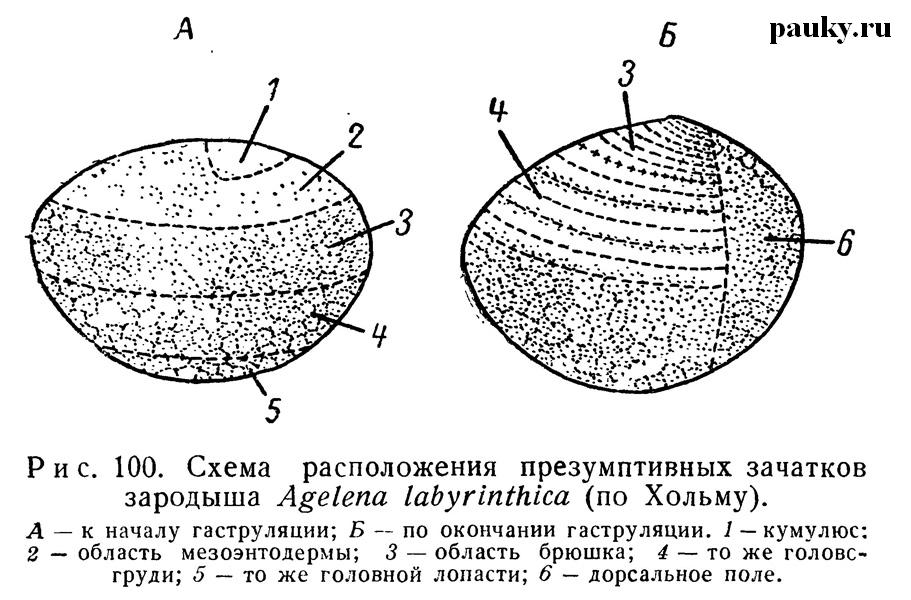
Вслед за тем область зародышевого диска занимается клетками будущей энтомезодермы, которые тоже мигрируют внутрь, после чего в области первичной полоски остается сзади эктодермальный материал будущего брюшка, а спереди — просомы. Расположение презумптивных зачатков энтодермы, энтомезодермы и эктодермы Хольм иллюстрировал специальными схемами (рис. 100).
Итак, к началу внешнего оформления зародышевой полоски под ее эктодермой уже имеются зачатки энто- и мезодермы. Позднее последняя образует пару мезодермальных полосок.
Разрозненные клетки, оставшиеся в желтке после обособления бластодермы, дают начало вителлофагам, перерабатывающим желток. Позднее они значительно пополняются за счет клеток, мигрирующих из мезоэнтодермального зачатка. Как и у других паукообразных, вителлофаги являются провизорными элементами и не участвуют в формировании средней кишки (Schimkewitsch, 1887); впрочем, по некоторым данным, часть их принимает участие в образовании печени.
Сегментация зародышевой полоски
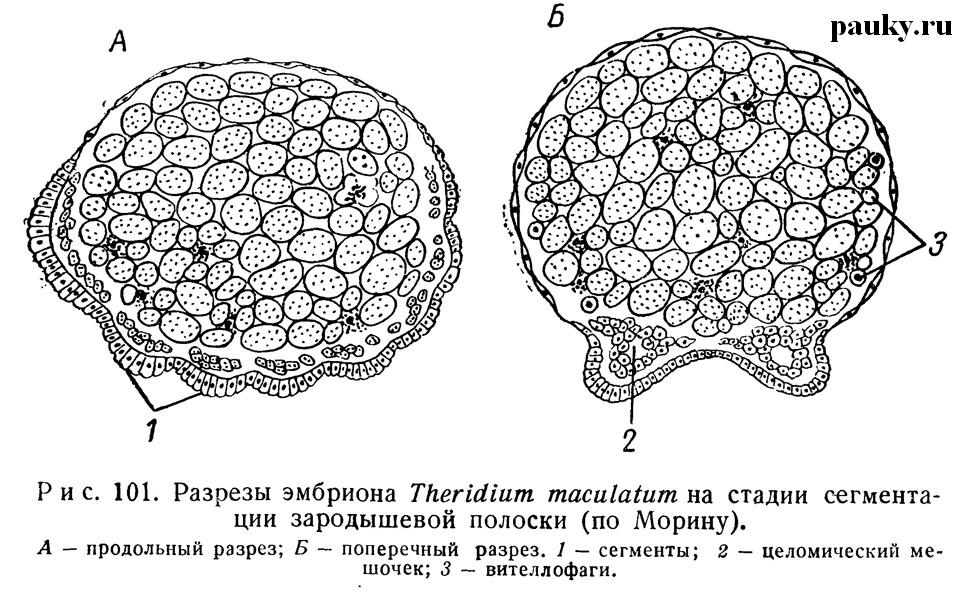
Мезодермальные полоски после их появления по сторонам срединной брюшной линии становятся двухслойными и разделяются на парные целомические мешочки (рис. 101), формирование которых тесно связано с внешним разделением зародышевой полоски на сегменты (см. рис. 99, Ж, 3).
По некоторым данным (Kishinouye, 1891; Pappenheim, 1903; Wallstabe, 1908), обособленные целомические мешочки появляются и в головной лопасти эмбриона (например, у Dolomedes и Agelena); некоторые рассматривают их как указание на наличие особого прехелицерального сегмента, другие считают вторичными образованиями. К этому времени зародышевая полоска настолько разрастается, что не только занимает всю брюшную поверхность яйца, но и заходит передними и задними концами на спинную сторону (см. рис. 99,3; рис. 102).
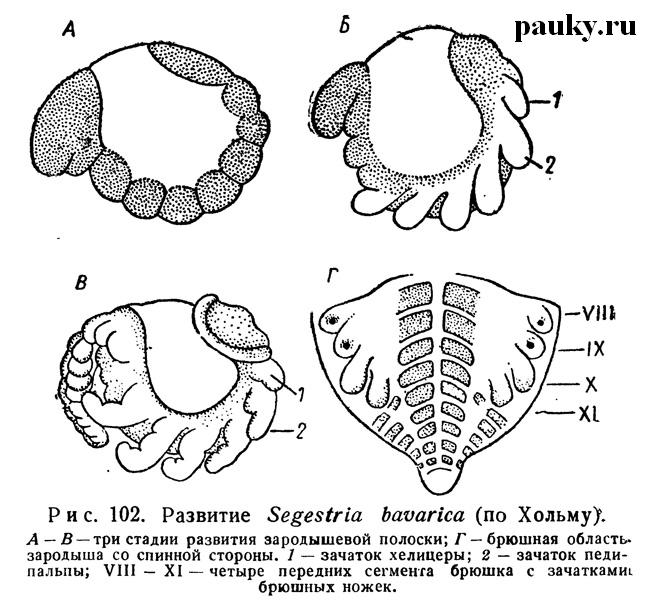
Сегментируясь, она разделяется на переднюю головную и заднюю анальную лопасти и несколько сегментов между ними. Сегменты принадлежат головогруди (просоме) и появляются не в порядке последовательности спереди назад, как у большинства паукообразных, но сперва обособляются четыре сегмента ходных ног и затем только впереди них формируются сначала сегмент педипальп, а потом и сегмент хелицер.
Однако, по старым наблюдениям Заленского (1871), у Clubiona прежде всего и притом одновременно дифференцируются лишь три сегмента просомы — сегменты педипальп, первой и второй пары ног, затем от анальной лопасти отчленяются самые задние сегменты просомы (третьей и четвертой пар ног), и только после этого из заднего края головной лопасти выделяется сегмент хелицер.
Трудно сказать, обнаружится ли такая же последовательность обособления сегментов головогруди и у других пауков при более глубоком изучении этого вопроса, но вряд ли могут быть сомнения, что в этом отношении Clubionidae чрезвычайно примитивны. Ранняя и одновременная закладка трех передних сегментов в данном случае не может быть объяснена иначе, как их ларвальной природой. Как показал П. Иванов (Iwanoff, 1933), у Chelicerata четыре передних сегмента являются ларвальными и гомологичны четырем протасписовым сегментам трилобитов. Позднее же обособление самого переднего из ларвальных сегментов — хелицерального характерно не только для всех пауков, но и для других паукообразных и представляет собой хотя и очень древнюю, но, несомненно, вторичную особенность.
Часто первичный порядок обособления ларвальных сегментов, в том виде как описано Заленским у Clubiona, утрачивается. В этом смысле большинство пауков, вероятно, являются вторично измененными. Но наиболее удаленными от исходных форм оказываются некоторые Entelegynae (Araneidae и Theridiidae), у которых зародышевая полоска остается узкой, а сегменты головогруди отчленяются последовательно от заднего к переднему, так что первым появляется сегмент четвертой пары ног, а последним хелицеральный.
Уже у Xiphosura самая передняя пара конечностей (развитие эмбриона паука) начинает запаздывать в своей закладке и дает маленькие хелицеры (P. Iwanoff, 1933). У Arachnida, и особенно у пауков, запаздывание четырех ларвальных сегментов настолько возрастает, что они начинают закладываться одновременно с передними постларвальными и, наконец, у некоторых Entelegynae даже отстают от них. То же по существу наблюдается и у насекомых: по мере их эволюции ларвальные, или первичноголовные, сегменты закладываются все позже, и у многих форм область грудных сегментов намечается уже на стадии бластодермы и даже до дробления (П. Иванов, 1937).





