
Большой сравнительно-анатомический интерес имеет дыхательная система пауков. Почти всегда она представлена двумя парами органов дыхания, принадлежащих VIII и IX туловищным сегментам. Лишь в редких случаях (Pholcidae, Symphytognathidae и некоторые другие отдельные формы) задняя пара редуцируется.
Легкие у пауков
Все примитивные группы обладают двумя парами легких. Таковы Liphistiomorphae, все Mygalomorphae и Hypochilidae из Araneomorphae. У громадного большинства аранеоморфных пауков на месте задней пары легких развиваются трахеи, и, наконец, Caponiidae, Telemidae и Symphytognathidae свойственны только трахеи. Таким образом, всех пауков можно разделить условно на три группы: четырехлегочных, двулегочных и безлегочных.
Число стигм (stigmata), или дыхалец, отвечает количеству органов дыхания, каждый орган имеет свою стигму, но у многих Araneomorphae задняя пара стигм, принадлежащая трахеям, сливается вторично, образуя непарное щелевидное дыхальце. (У Salticus, например, сперва закладываются две стигмы трахей, которые затем сливаются.)
Несомненно, более древним типом дыхательных органов являются легкие. Несмотря на недостаточную изученность их в разных группах, совершенно ясно, что они отличаются большим постоянством строения.
Стигмы легких имеют вид поперечных щелевидных отверстий. Каждая стигма открывается в так называемое преддверие (atrium, vestibulum — рис. 56) легкого, которое в то же время является задним отделом легочного мешка.
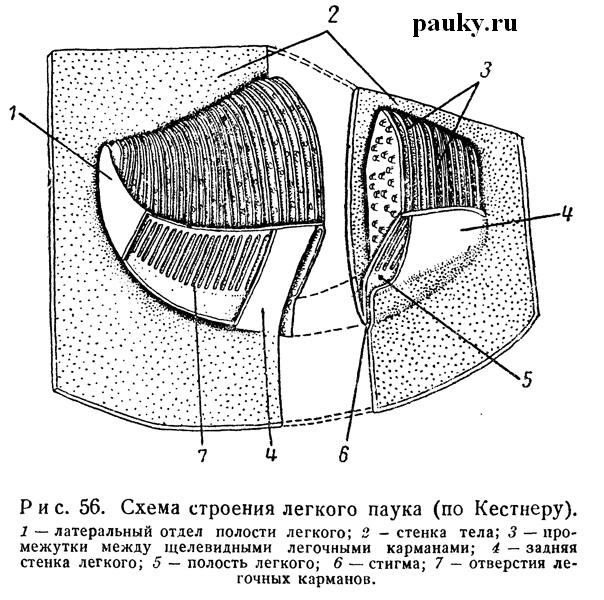
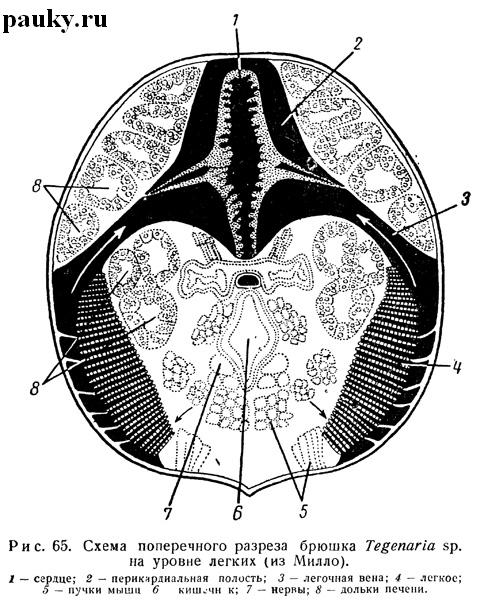
Последний всегда лежит впереди своей стигмы, уплощен дорсо-вентрально, особенно спереди, и имеет более или менее округлую или, чаще, треугольную форму. Легкие окружены окололегочными кровеносными синусами (см. рис. 65), передние концы которых, суживаясь, образуют легочные вены.
По общему плану строения легкие пауков очень напоминают таковые Scorpiones, Uropygi и Amblypygi. Большая, передняя часть легочного мешка состоит из многочисленных легочных пластинок, задняя является преддверием. Пластинки представляют собой тонкие уплощенные складки стенок легкого (рис. 56). Они обычно имеют форму вытянутых равнобедренных треугольников, обращенных коротким основанием назад, и лежат параллельно друг другу и почти параллельно брюшной стенке тела. Края их сращены со стенками легкого, за исключением задних краев, обращенных в сторону преддверия. Между последними лежат многочисленные параллельные щели, ведущие в узкие промежутки между легочными пластинками.
Каждая пластинка ограничена очень тонкой хитиновой кутикулой (рис. 57), которая, однако, на заднем свободном крае пластинки значительно утолщается.
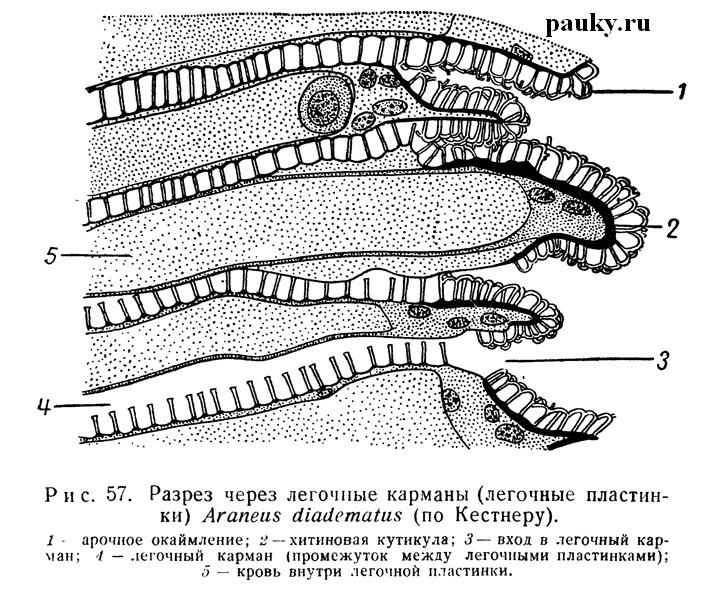
На верхней стороне пластинки несут маленькие хитиновые столбики, слегка расширенные на концах; они не позволяют соседним пластинкам слипаться, благодаря чему между пластинками всегда имеется воздух. На задних краях пластинок концы столбиков соединяются друг с другом дужками, образующими характерное арочное окаймление. Хитиновые столбики развиты обычно и на задней стенке преддверия. Под кутикулой пластинок лежит тонкая гиподерма, утолщающаяся в области заднего края. Узкие лакунарные пространства легочных пластинок пересекаются соединительнотканными опорными мостиками, соединяющими обе стенки. Эти пространства наполнены кровью, проникающей сюда из окололегочного синуса. Газообмен совершается через тонкие стенки пластинок. Легкое прочно фиксируется на брюшной стенке тела посредством мелких мускульных пучков, прикрепляющихся к глубоким складкам хитина легочной крышечки.
Описанное строение имеют легкие паука-крестовика (Araneus diadematus), но отличия, встречающиеся у других форм, сводятся лишь к второстепенным деталям. Значительно колеблется число легочных пластинок не только у разных видов (у Liphtstius malayanus— 140—150, у Segestria senoculata — 25, у самки Araneus diadematus — более 100), но даже у особей одного вида, в зависимости от возраста и пола. Несомненно, существует и коррелятивная зависимость числа легочных пластинок от размеров животного и степени развития трахей.
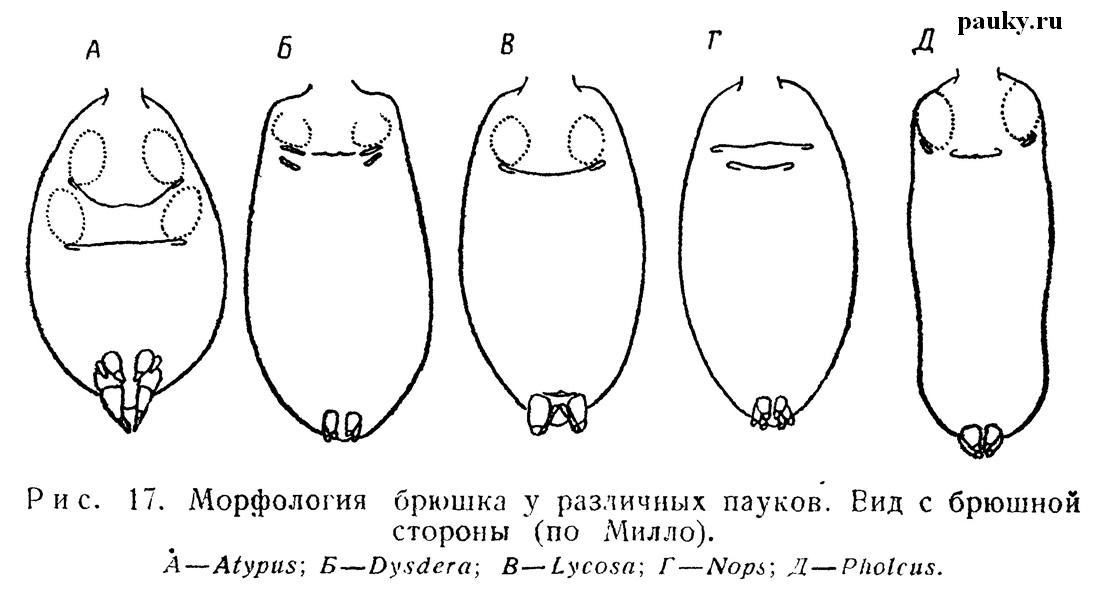
Первая пара легочных стигм у Liphistiomorphae и стигмы единственной пары легких у Entelegynae тесно связаны с эпигастральной бороздой. Стигмы здесь представляют собой всего лишь боковые расширения эпигастральной щели (см. рис. 17,А, Б, В, Д).
Соответственно и преддверия легких на внутренних своих краях соединяются с узкой полостью эпигастральной складки, а через нее — друг с другом. Половое преддверие и аподемы эндостернитов VIII сегмента, представляющие собой участки эпигастральной складки, связаны, таким образом, у этих пауков с преддвериями легких (см. рис. 43).
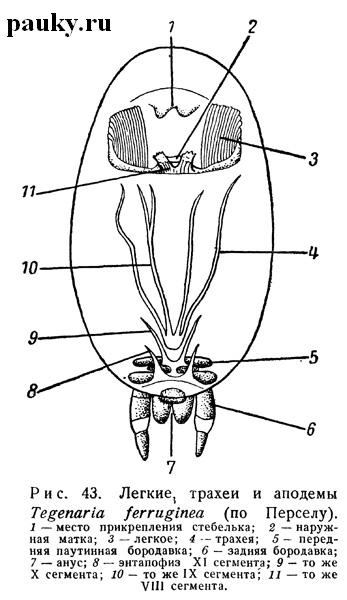
У Mygalomorphae и части Araneomorphae эти же стигмы независимы от эпигастральной борозды, хотя и лежат на одном уровне с половым отверстием и точками прикрепления эндостернитов.

Трахеи пауков
Трахеи, свойственные только Araneomorphae, отличаются большим разнообразием. Как сказано, у двулегочных пауков они развиваются на месте второй пары легких, т. е. в IX сегменте, но в трех семействах — Telemidae, Caponiidae и Symphytognathidae — первая пара легких замещена трахеями.
У этих безлегочных пауков первая пара трахеальных стигм расположена в тех же самых местах, где у других форм лежат стигмы первой пары легких. Более того, у Caponiidae эти стигмы суть лишь боковые участки эпигастральной борозды, подобно первым стигмам Liphistiidae и Entelegynae, а у Symphytognathidae основания трахей соединены друг с другом поперечным трахеальным стволом, который, по-видимому, является производным средней части эпигастральной складки.
Вторая пара стигм у пауков без легких может быть расположена на большем удалении от первой пары (Telemidae), но чаще помещается значительно ближе к ней, чем соответственные стигмы у четырехлегочных форм (Caponiidae — см. рис. 17,Г), или, наконец, редуцируются вовсе (Syfnphytognathidae).
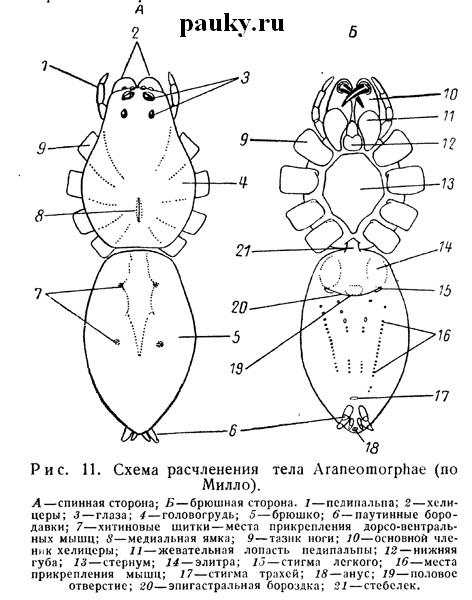
У части двулегочных пауков (Oonopidae и Dysderidae) трахейные стигмы помещаются близко позади легочных (рис. 17,Б). По-видимому, с заменой одного типа дыхательных органов другим стигмы сдвигаются вперед, хотя и остаются на том же сегменте. У остальных же двулегочных трахейные стигмы сливаются в непарное щелевидное дыхальце. Оно остается недалеко от эпигастральной борозды лишь у немногих форм, но у большинства благодаря разрастанию IX сегмента смещаются далеко назад на его границу, помещаясь впереди паутинных бородавок (см. рис. 11, 17,В).
Что касается самих трахей, то по степени развития и физиологическим особенностям они могут быть разделены на два типа, между которыми имеются переходы. Различают трахеи местного и общего значения. Первые не распространяются за пределы своего сегмента или во всяком случае ограничены областью брюшка. Вторые проникают сквозь стебелек в головогрудь и в конечности
Три семейства безлегочных пауков обнаруживают интересные примеры корреляции между трахеями первой и второй пары и приспособления к равномерному снабжению кислородом всех органов тела. Так, у Caponiidae (рис. 58) каждая передняя стигма ведет в мешковидное преддверие, от которого отходит большое количество очень тонких и коротких трахейных трубочек.
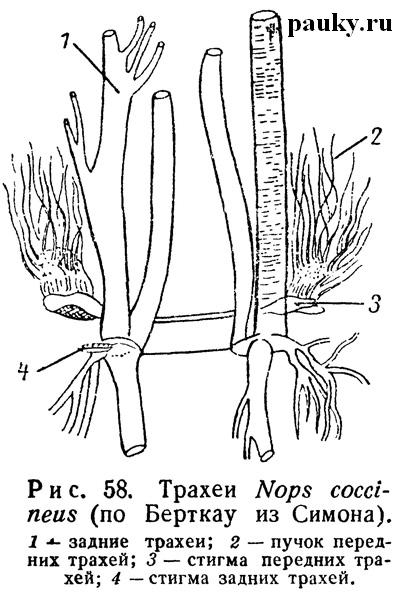
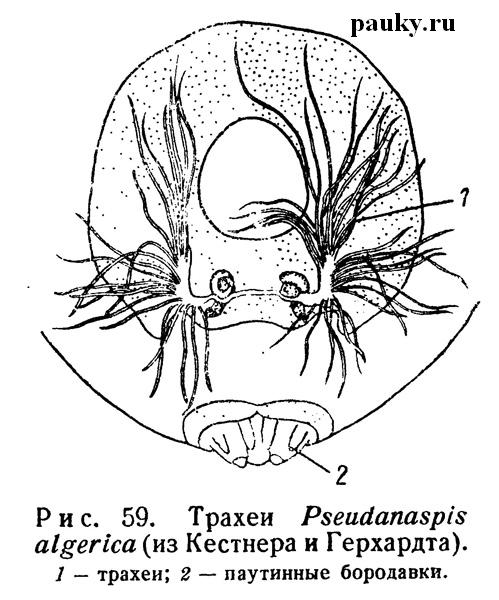
Омываясь циркумтрахеальным кровеносным синусом, они обогащают гемолимфу кислородом, т. е. функционируют подобно легким. Трахеи же задней пары берут на себя доставку воздуха непосредственно к месту потребления кислорода. От каждой задней стигмы отходит несколько толстых трубчатых стволов. Из них часть разветвляется в брюшке, другая проникает через стебелек в головогрудь и здесь дает ветви к мускулатуре и во все конечности. У Symphytognatidae (рис. 59) единственная пара передних стигм ведет в небольшие преддверия, от которых начинается множество длинных трахей, снабжающих как брюшко, так и головогрудь, где они проникают в конечности.
Наконец, у Telemidae трахеи передних стигм (четыре ствола на каждой стороне) обслуживают почти исключительно органы головогруди, тогда как трахеи вторых стигм (5—8 пар стволов) снабжают воздухом только брюшко.
Переходя к трахеям двулегочных пауков, рассмотрим сперва формы с трахейными стигмами, лежащими непосредственно позади легочных стигм. Весьма своеобразны трахеи Dysderidae (рис. 60).
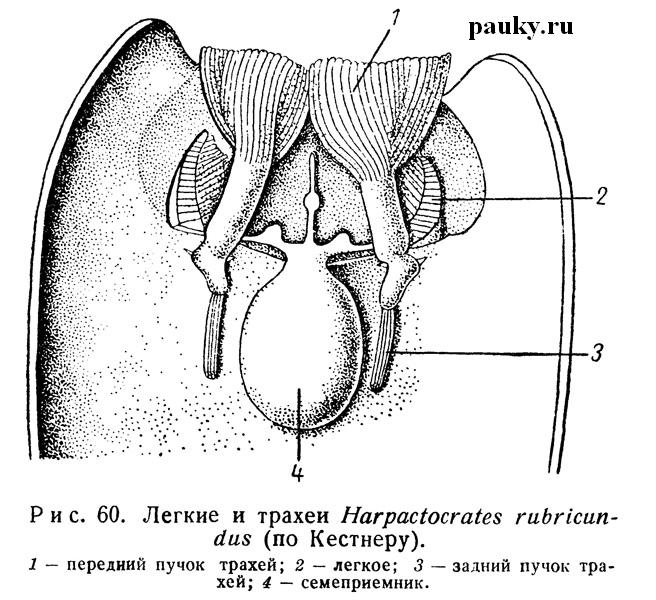
Каждая стигма открывается в короткий и широкий трахейный ствол, от задней части которого отходит назад густой пучок тонких капиллярных трубок, распределяющих воздух в пределах брюшка. Передние концы стволов дают по густому мощному пучку таких же трубок, проходящих в головогрудь, где они оплетают кишечник, мозг и заходят в конечности. Сходного строения трахеи свойственны и Oonopidae, но, в отличие от Dysderidae, стигмы у них связаны интертрахеальной складкой покровов. Последняя является задней пограничной складкой стенки тела IX сегмента, т. е. аналогична эпигастральной борозде на VIII сегменте.
Но особенно отчетлива связь второй пары трахей с задней пограничной складкой сегмента у тех пауков, у которых единственная трахейная стигма более или менее сдвинута назад. Так, у Filistatidae щелевидная стигма открывается в полость складки. От последней отходят две пары коротких полых трубочек (рис. 61), причем средняя пара, несомненно, является аподемами IX сегмента, так как к ней прилегают эндостерниты этого сегмента. Обе пары трубок функционируют как трахеи.
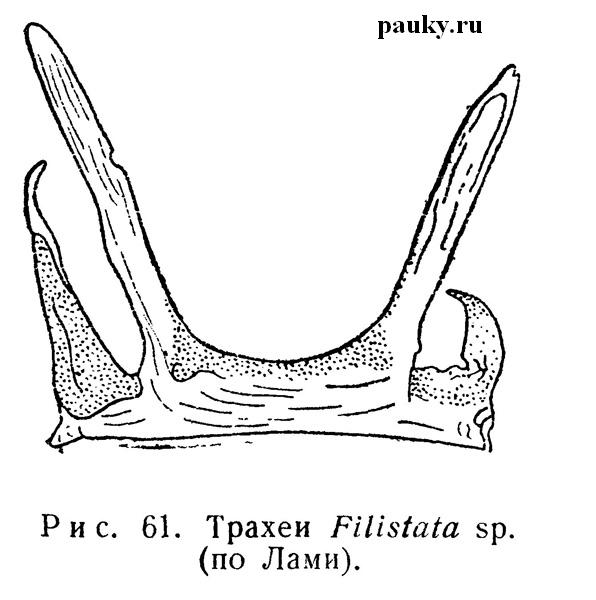
Кутикула в глубине складки и в начале аподем покрыта хитиновыми палочками, напоминающими таковые на пластинках легкого. Считается, что трахеи Filistatidae находятся на начальной ступени формирования, хотя некоторые авторы (Petrunkevitch, 1928) рассматривают их как редуцирующиеся органы.
У других двулегочных пауков с непарной стигмой обе пары придатков развиваются в настоящие трахеи, которые остаются простыми неразветвленными трубками или могут, так или иначе, разветвляться.
Громадное большинство Araneomorphae (Araneidae, Lycosidae, Clubionidae, Agelenidae и мн. др.) как раз характеризуется наличием четырех трахейных стволов, в виде длинных неразветвленных трубок (см. рис. 43). Наружная пара, как показывает история развития, формируется на месте второй пары легких. Средняя пара даже у взрослых особей связана с эндостернитами IX сегмента, что не оставляет никаких сомнений в ее гомологии аподемальным впячиваниям.
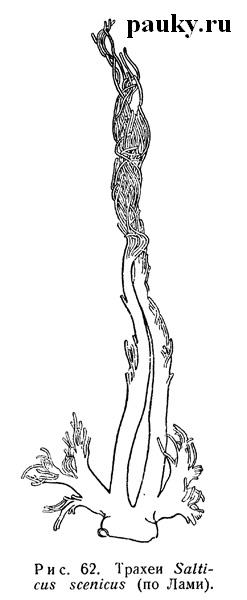 Дальнейшее усложнение трахей проявляется в развитии на них концевых разветвлений в виде многочисленных капиллярных трубочек (Salticidae — рис. 62, Uloboridae, Thomisidae и др.), причем разветвления средней пары проникают через стебелек в головогрудь, доставляя воздух в мускулатуру, конечности и другие органы. Замечательно, что даже при таком мощном развитии связь средних трахей с эндостернитами IX сегмента сохраняется.
Дальнейшее усложнение трахей проявляется в развитии на них концевых разветвлений в виде многочисленных капиллярных трубочек (Salticidae — рис. 62, Uloboridae, Thomisidae и др.), причем разветвления средней пары проникают через стебелек в головогрудь, доставляя воздух в мускулатуру, конечности и другие органы. Замечательно, что даже при таком мощном развитии связь средних трахей с эндостернитами IX сегмента сохраняется.
Существует, кроме того, целый ряд форм, у которых развиваются только эти аподемальные по происхождению трахеи (Dietynidae, некоторые Agelenidae, Argyronetidae). Разветвления их лежат частью в брюшке, частью в головогруди.
Наконец, полная редукция трахей свойственна Pholcidae, Diguetia и одному чилийскому виду Sicarius. У этих пауков остается только одна пара легких.
Гистологическое строение трахей имеет некоторые особенности. Под тонкой кутикулой лежит слой гиподермы с эллиптическими ядрами. Кутикула несет многочисленные хитиновые палочки, простые или несколько сжатые на конце. Палочки у своей вершины иногда дают перпендикулярные ветви, утолщенные у основания. Если они анастомозируют друг с другом, то над кутикулой образуется нежная ситовидно продырявленная пластинка (Dysdera и др.). Все эти кутикулярные образования очень похожи на палочковый аппарат легких и служат той же цели, препятствуя спадению просвета трахей. Дальнейшая дифференциация проявляется в развитии спирального опорного утолщения кутикулы, которое имеется в трахеях многих пауков (Uloboridae, Dysderidae) и так характерно для сольпуг и насекомых.
В табл. 3 представлены все главные типы органов дыхания, известные у пауков.

Физиология дыхания у пауков
Обновление воздуха в легких происходит благодаря вентиляции, осуществляемой работой особых мускульных пучков — открывателей стигм. Эти мышцы спереди прикрепляются к задней стенке преддверия легкого над краем стигмы и тянутся назад и внутрь к брюшной стенке тела. Сокращаясь, они тянут назад заднюю стенку преддверия и открывают стигмы. При этом полость преддверия резко расширяется, и в него заходит воздух извне. При расслаблении мускула задняя стенка преддверия, вследствие своей эластичности, занимает прежнее положение, объем преддверия уменьшается, и часть воздуха выходит наружу, а стигма закрывается.

Обновление воздуха в узких пространствах между легочными пластинками совершается главным образом путем диффузии.
Значение легочного дыхания у пауков выяснено экспериментами над Tegenaria и Lycosidae. Если у этих пауков смазать вазелином стигмы легких, то уже через 2—3 минуты они сильно слабеют, частота сердечных сокращений быстро падает, и через 4—7 часов невозможно уже заметить никаких следов кровообращения. Выключение легких приводит к параличу дыхательных центров и через несколько часов — к смерти. Сравнительно слабо развитые трахеи, таким образом, не могут заменить легких, и, очевидно, роль их в дыхании незначительна. Совсем другие результаты дали опыты над Segestria senoculata — пауком с богато разветвленными, заходящими в головогрудь трахеями. Замазывание легких не производит на него заметного действия, и ясно, что газообмен легко совершается через трахеи.
По-видимому, увеличение количества углекислоты в легких стимулирует открывание стигм. Паук, погруженный на некоторый срок в воду, широко раскрывает легочные стигмы. То же самое наблюдается, если заставить животное некоторое время быстро бегать.

Интересно, что у водяного паука — Argyroneta aquatica преобладающее значение имеет кожное дыхание, а роль трахей и легких в дыхании сравнительно невелика. Животное с замазанными вазелином стигмами живет от 4 до 15 дней и погибает затем от избытка углекислоты в крови. Под водой водяной паук дышит за счет воздуха, облекающего его тело серебристой мантией. Поэтому, подобно многим водным насекомым, время от времени он поднимается на поверхность водоема для обновления своего воздушного запаса. Высунув из воды кончик брюшка и провентилировав легкие и трахеи, паук погружается в воду с новым запасом воздуха на поверхности стернума и брюшка. Здесь воздух удерживается в густом покрове гидрофобных слегка перистых волосков. На брюшке, где рни особенно сильно развиты, имеется около 1200, а на стернальной пластинке — да 750 волосков на 1 мм2. Воздух остается между волосками благодаря, их густоте и несмачиваемости. Среди них различаются два сорта — многочисленные короткие и более длинные и жесткие. Последние подпирают пленку натяжения между слоем воздуха и водой. У самцов, в отличие от самок, часть спинной поверхности брюшка лишена волосков. Поэтому и форма воздушной мантии у них несколько отлична. От времени до времени паук расчесывает и смазывает волосяной покров кончиками четвертой и третьей пар ног. Лапки и пятки ног при этом сначала протягиваются между хелицерами и смачиваются прозрачным секретом, выступающим из предротовой полости. Этот своеобразный «туалет», предупреждающий гибельное для паука развитие на волосках бактерий, совершается им в воздушном колоколе.

Значительное время Argyroneta проводит в подводном гнезде — воздушном колоколе, где отдыхает, подстерегает добычу поедает ее и где происходит копуляция. Как показали наблюдения в лаборатории, паук может оставаться в колоколе до трех недель, не возобновляя его воздушного запаса. Летом воздух колокола содержит от 5 до 10% кислорода, в зависимости от длительности пребывания в нем паука. Однако через некоторое время между воздухом колокола и водой происходит обмен газами — воздух несколько обогащается кислородом и теряет углекислоту. Потребление кислорода взрослой самкой в течение часа при температуре 18° достигает 7 мм3, а при 2° падает до 0,7 мм3 (Braun, 1931).





