
Все пауки раздельнополы; гермафродитизм встречается только как ненормальное явление и крайне редко.
Половой диморфизм
Различие между полами является правилом, из которого, однако, имеются исключения. Более или менее одинаковые размеры и внешность самца и самки свойственны многим формам из Mygalomorphae, Gnaphosidae, Pholcidae и Lycosidae. Иногда половой диморфизм выражается в удлинении ног у самцов (например, у Clubionidae), причем эта особенность проявляется у них только после последней линьки и связана с их более подвижным образом жизни пауков.
Сплошь и рядом самец по размерам значительно уступает самке (Thomisidae, Salticidae и мн. др.). Особенно Мелки самцы у Araneidae. Сравнительно слабые различия между полами у Araneus возрастают у Argiope, Gasteracantha, Micrathena, Cyrtophora и особенно у Nephila. У некоторых видов Nephila самка по объему превышает в 1000—1500 раз самца, который вполне заслуживает название карликового. Для многих видов в родах, характеризующихся мелкими самцами, до сих пор известны лишь самки, что объясняется отнюдь не редкостью самцов, но скрытным их образом жизни и недолговечностью.
У Eresus niger (Cribellatae, Eresidae) описан диморфизм самцов. Самка достигает 30 мм в длину, а среди самцов имеются крупные особи в 16—17 мм и мелкие в 10—12 мм, причем различия между двумя формами самцов не ограничиваются размерами тела (Сычевская, 1954).
Обратные отношения, когда самец крупнее самки, очень редки (Argyroneta aquatica и Harpactes hombergi).
Помимо того, часто половой диморфизм выражается в тех или иных вторичных половых признаках. Нередко самцы отличаются окраской. Так, например, среди Sparassidae самец Micrommata virescens отмечен на спине красивой ярко-красной полоской, тогда как самка однообразно зеленоватая. У Eresus niger самец отличается от скромно окрашенной самки черно-красной расцветкой. Salticidae свойственны еще более резкие различия между полами.
По внешнему виду и орнаментации самец нередко не имеет никакого сходства с самкой. У самца Saitis barbipes третья пара ног удлинена и украшена пучками длинных волосков, а тело несет красивый рисунок, образованный разноцветными волосками, тогда как самка сохраняет скромную окраску. В киноварно-красный цвет окрашена почти вся верхняя сторона и бока брюшка у самца Philaeus chrysops, в отличие от черной самки.
В целом ряде случаев самцы и самки различаются формой головогруди или брюшка, что особенно характерно для некоторых Linyphiidae и Araneidae.
Особый интерес представляют органы самцов, служащие для удерживания самки при копуляции. Самцы многих Mygalomorphae имеют большой вырост на голенях первой пары ног; во время спаривания между этими выростами и метатарзальными члениками зажимаются ноги самки. Сходный вырост на голенях имеет и самец паука-крестовика, а самец паука Tetragnatha — на хелицерах. Наконец, массивные и необычайно вытянутые хелицеры самца Dictyna тоже приспособлены для удерживания самки: самец зажимает между их внутренними краями, снабженными специальными выемками, хелицеры самки.
У некоторых форм самцы имеют органы стридуляции (Asagena, Steatoda). Наконец, зрелые самцы Cribellatae лишены крибеллум и каламиструм, что связано с особенностями их биологии.
Мужская половая система
Внутренние органы половой системы просты. Компактные и парные, как правило, гонады помещаются в брюшке под печенью и над паутинными железами. Спереди от них отходят два половых протока, которые затем соединяются, образуя непарный концевой канал. Единственное простое половое отверстие представляет собой медиальную часть эпигастральной щели между стигмами передних дыхательных органов.
Семенники (рис. 81) имеют вытянутую, цилиндрическую форму и нередко образуют извивы. У Mygalomorphae и Dysderidae их задние концы сливаются, как у сенокосцев и клещей, поэтому гонада непарная. У многих других форм кажущееся сходство с таким подковообразным семенником вызвано тесной спайкой задних концов гонады посредством богатой жиром соединительной ткани.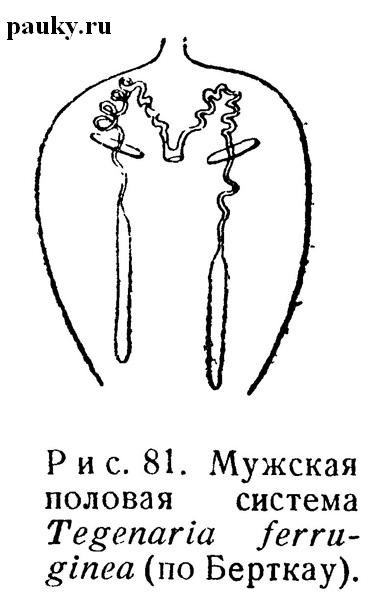
Семяпроводы (рис. 81) в большинстве случаев резко отграничены от семенников и более или менее извиты. Непарный конечный проток иногда образует двулопастной мешочек, называемый мужской маткой (uterus masculinus) (Schimkewitsch, 1884).
Сперматогенез пауков, известен главным образом благодаря исследованиям Ю. Вагнера (1896),: Безенберга (Bosenberg, 1905), Пайнтера (Painter, 1914) и И. И. Соколова (1960). По данным И. И. Соколова, гаплоидное число хромосом у пауков, обнаруживающееся во время сперматогенеза, колеблется от 8 до 24 (чаще в пределах от 12 до 15). И. И. Соколов выяснил кариологические особенности сперматогенеза и проследил также непосредственный переход нитевидных хромосом ядра сперматиды в формирующуюся головку спермия, в которой они остаются видимыми и в дальнейшем. Это наблюдение имеет большой интерес, так как обычно ядро спермия считается бесструктурным.
Сперматозоиды пауков заключены в особые микроскопически малые мешочки — капсулы, которые, однако, не являются настоящими сперматофорами. У примитивных форм (все Mygalomorphae), а также у Dysderidae и Oonopidae это так называемые ценоспермии. Количество сперматозоидов, находящихся в их округлых капсулах, варьирует весьма широко даже в пределах одного рода. Так, у Segestria senoculata их 4, а у S. bavarica — от 60 до 100. Семенная жидкость высших пауков Entelegynae содержит клейстоспермии, оболочки которых заключают только по одному сперматозоиду.
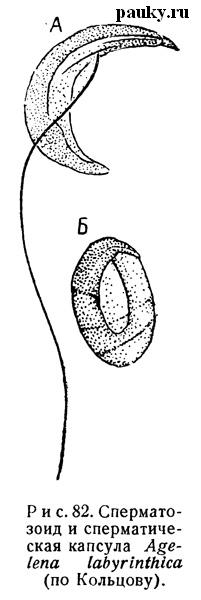 Еще Ю. Вагнер (1896) показал, что спермий Entelegynae своеобразно видоизменяются во время спермиогенеза. В конце своего формирования они имеют обычное строение: обладают длинной цилиндрической головкой с перфораторием, продольной и двумя спиральными скелетными нитями, подвижной хвостовой нитью и на месте шейки — протоплазматическим пузырьком. При дальнейшем развитии, однако, шейка не формируется, пузырек не отрывается, как обычно, но головка и хвостовая нить обвертываются вокруг него. Наконец, весь спермий превращается в яйцевидное или шарообразное тельце, одетое твердой капсулой (рис. 82). По Н. К. Кольцову (1936), такое своеобразное превращение жгутикового спермия является настоящим инцистированием. Оболочка выдерживает кипячение в щелочах и, по-видимому, состоит из хитиноидного вещества. В семяпроводы поступают только инкапсулированные спермин.
Еще Ю. Вагнер (1896) показал, что спермий Entelegynae своеобразно видоизменяются во время спермиогенеза. В конце своего формирования они имеют обычное строение: обладают длинной цилиндрической головкой с перфораторием, продольной и двумя спиральными скелетными нитями, подвижной хвостовой нитью и на месте шейки — протоплазматическим пузырьком. При дальнейшем развитии, однако, шейка не формируется, пузырек не отрывается, как обычно, но головка и хвостовая нить обвертываются вокруг него. Наконец, весь спермий превращается в яйцевидное или шарообразное тельце, одетое твердой капсулой (рис. 82). По Н. К. Кольцову (1936), такое своеобразное превращение жгутикового спермия является настоящим инцистированием. Оболочка выдерживает кипячение в щелочах и, по-видимому, состоит из хитиноидного вещества. В семяпроводы поступают только инкапсулированные спермин.
В свежевыделенной сперме сперматозоиды неподвижны. Но при разбавлении семенной жидкости водой они, выходя из капсул, начинают оживленно двигаться. По наблюдениям Н. К. Кольцова, тоже самое происходит, когда сперма попадает в семеприемники самки после спаривания. Здесь капсулы лопаются, и спермий благодаря эластичности их скелетных нитей распрямляются, принимают обычную форму и начинают оживленно двигаться при помощи хвостика.
Биологическое значение инцистирования спермиев, свойственного, помимо пауков, еще и псевдоскорпионам, было разъяснено И. И. Соколовым (1926). Спаривание в обеих группах протекает различно, но имеется один общий момент: маленькая капелька спермы, выделенная самцом (совокупительный аппарат самцов паука), остается некоторое время на воздухе. У псевдоскорпионов это происходит непосредственно во время спаривания, у пауков — до него, перед наполнением самцом педипальпальных совокупительных органов, когда сперма выделяется на специально сплетаемую паутиновую сеточку.
Капсулы, в которые заключены спермии, очевидно, являются приспособлением, предохраняющим их от высыхания и от других вредных воздействий воздушной среды.
Кроме капсул, семенная жидкость содержит массу мелких зерен, которым приписывается питательная роль по отношению к спермиям. Их происхождение неясно; вероятно, они образуются путем распадения плазматических остатков при сперматогенезе или абортивных сперматогониев и сперматоцитов.
Женская половая система
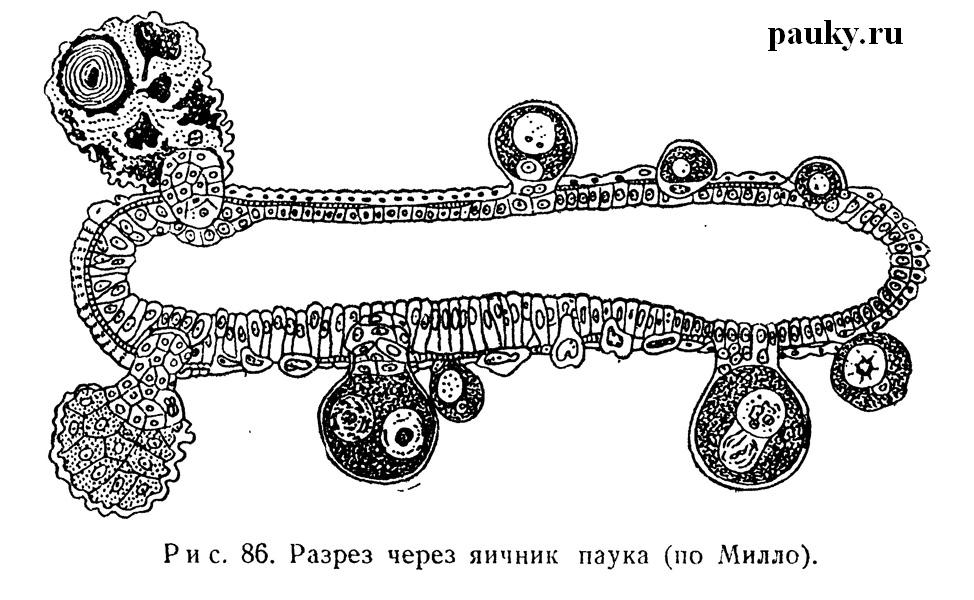
Яичники у самок пауков в большинстве случаев мешковидны (рис. 85). Только у Mygalomorphae и Dysderidae описан непарный подковообразный яичник, образующийся, как и непарный семенник этих форм, путем срастания концов парной гонады. Строение яичников было изучено у Araneus diadematus Тревиранусом (Treviranus, 1812) и Шимкевичем (Schimkewitsch, 1884). Однослойный зачатковый эпителий его стенок ограничен снаружи перитонеумом. Ооциты выклиниваются из зачаткового слоя на наружную поверхность органа.
Здесь они остаются до созревания, будучи связаны с эпителием яичника особым клеточным стебельком (рис. 86). Зрелые яйца уходят во внутреннюю полость яичника, откуда уже выводятся, наружу по яйцеводам. Подобное же строекие яичников характерно для всех Chelicerata.
После ухода яйца внутрь стебелек обогащается липоидными включениями и образует на поверхности яичника пятнышко, которое сравнивают с желтым телом млекопитающих. Эпителиальные клетки зачаткового эпителия, не развивающиеся в ооциты, приобретают железистый характер. В момент кладки яиц они выделяют в изобилии зернистый, ацидофильный секрет, сходный по составу с фиброином шелка (Balbiani, 1897). Выходя вместе с яйцами наружу, этот секрет составляет цемент, слегка склеивающий яйца.
Парные яйцеводы (см. рис. 85) впадают в сходный с ними по гистологии и тоже мезодермальный по происхождению непарный канал внутренней матки (uterus internus). Продолжение последней — наружная матка (u. externus), ИЛИ влагалище (vagina), имеет эктодермальную природу, выстлано хитином и открывается первичным половым отверстием непосредственно, наружу (Liphistiomorphae, Mygalomorphae, Haplogynae) или в глубине эпигастральной складки (Entelegynae). В последнем случае яйца откладываются через медиальный участок эпигастральной щели, который, следовательно, играет роль вторичного полового отверстия (см. рис. 15, Б).
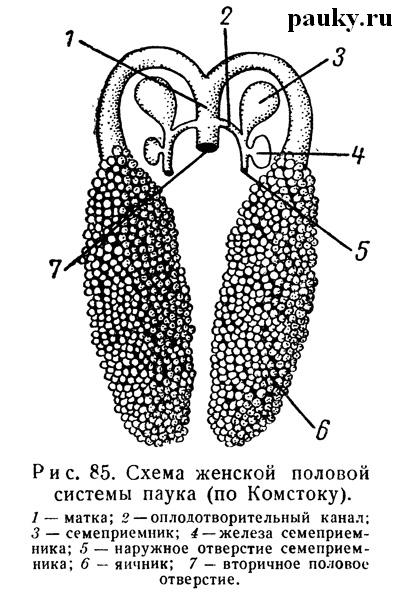
В связи с половым отверстием самки находятся совокупительные органы, т. е. семеприемники (receptacula seminis — рис. 87) и их каналы. У Aviculariidae, Haplogynae и Tetragnatha семеприемники открываются во влагалище и являются дериватами последнего, тогда как у всех остальных пауков отверстия их самостоятельны. Чаще всего бывает 2 семеприемника, но у Tetragnatha их 3, у Segestria — 5, Clubiona — 6, а у Aiypus — даже 28.
Каждый представляет собой округлый мешочек с каналом, по которому при спаривании поступает сперма. У громадного большинства пауков из семеприемников сперма проникает по оплодотворительным каналам во влагалище, где происходит оплодотворение (рис. 87). Семеприемники и связанные с ними каналы выстланы хитином. Гиподерма семеприемников часто содержит железистые клетки, открывающиеся в его полость мелкими порами. Усиление железистой функции семеприемников у ряда пауков (Agelena, Tibellus oblongus) приводит к превращению их в чисто железистые образования (Engelhard, 1910).
У Entelegynae наружные отверстия каналов семеприемников лежат на хитинизированной площадке эпигины (epigyne — рис. 87) и служат для введения в них совокупительных органов самца. Влагалище, таким образом, не оправдывает своего названия. Параллельно с усложнением органов совокупления у самцов усложняется и строение каналов семеприемников.
У некоторых форм эпигина несет более или менее длинный и гибкий придаток (clavus — рис. 87,А). Прежде считалось, что он служит яйцекладом. В действительности же он играет какую-то роль при спаривании.





