
Сильно концентрированная центральная нервная система пауков целиком сосредоточена в головогруди. В этом отношении пауки сходны с Pseudoscorpiones, Opiliones и Acari. Встречающиеся до сих пор в литературе указания на то, что у низших пауков задний, сложный по составу ганглий брюшной нервной цепочки лежит обособленно в брюшке (сходно с Uropygi и Solifugae) неверны. Центральная нервная система состоит из головного мозга и подглоточной нервной массы (рис. 70).
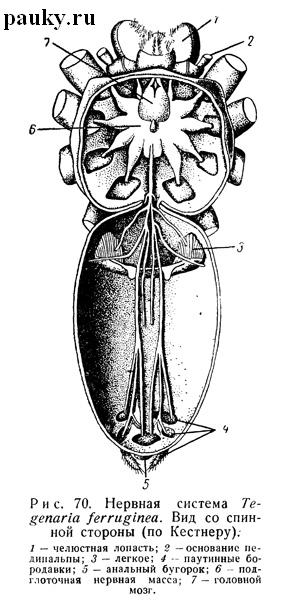
Последняя помещается на брюшной стороне между стернальной пластинкой или слепыми выростами желудка и эндостернитом.
Головной мозг у пауков
Головной мозг (см. рис. 70, 72) расположен над пищеводом. Он сравнительно невелик и обычно несет на верхней и передней стороне медиальную бороздку.
С подглоточной нервной массой мозг соединяется по бокам от пищевода. Как у всех Chelicerata, мозг состоит только из протоцеребрума и тритоцеребрума. От второго отдела мозга — дейтоцеребрума, дающего у Crustacea и Tracheata нервы к антеннам, у пауков не остается следа. Нет, так же как и у других паукообразных, внешнего разделения между прото и тритоцеребрумом. Мозг иннервирует глаза и хелицеры и дает начало стомато-гастрической нервной системе.
Гистология мозга довольно хорошо изучена (Hanstrom, 1919, 1935). В отличие от других паукообразных, передняя сторона мозга пауков глубоко вдавлена внутрь (рис. 71). Поэтому основания главных нервов сближены друг с другом медиально.
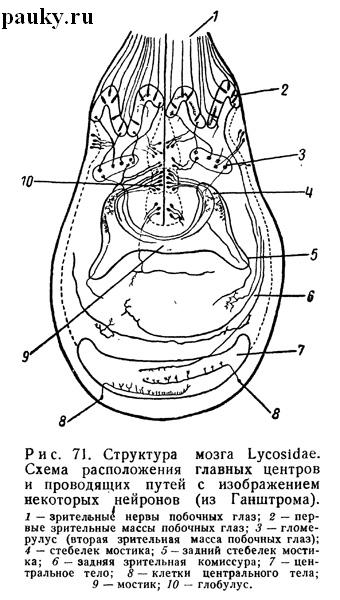
Как обычно, периферический слой мозга образован многочисленными ганглиозными клетками, а центральная область занята густым сплетением нервных волокон. В состав прото-церебрума входят зрительные ганглии с проводящими зрительными путями (комиссурами), большое центральное тело (corpus centrale) и пара грибовидных тел. Густая волокнистая масса центрального тела состоит из отростков мелких нервных клеток, образующих полулунную мантию на задней поверхности мозга.
Древовидно разветвленные волокна центрального тела соединяются с отростками нервных клеток других частей мозга, и, очевидно, этот орган является главным ассоциативным центром. Такое же значение имеют и грибовидные тела. Каждое состоит из шапочки нервных клеток, отростки которых образуют волокнистый стебелек, связанный с парным ему стебельком противоположной стороны мощным волокнистым мостиком. Однако в состав последнего входят и волокна от других частей мозга. Особенное значение придается многочисленным волокнам, проникающим в мостик от зрительных ганглиев.
Имеются два парных зрительных центра. Один связан с так называемыми главными глазами (передними медиальными), другой — со всеми остальными.
Это небольшие сгущения нервных волокон, каждое со своей группой мелких зрительных клеток в периферической коре мозга. Парный центр главных глаз лежит в теменной части мозга. Он состоит на каждой стороне из двух зрительных ганглиев. Из них передний принимает окончания зрительных волокон сетчатки и связан волокнистым пучком со вторым, лежащим несколько сзади. Длинная дугообразная зрительная (задняя), комиссура соединяет вторые зрительные ганглии друг с другом и дает волокна в центральное тело. Вместе со зрительными ганглиями главных глаз эта комиссура образует так называемый верхний зрительный путь.

Парные зрительные центры остальных глаз, связанные подобной же дугообразной комиссурой, принадлежат к нижнему зрительному пути. Их зрительные ганглии на каждой стороне распадаются на три части соответственно трем зрительным нервам. Они принимают нервные отростки зрительных клеток сетчатки и связаны волокнами со зрительной комиссурой, вторыми зрительными ганглиями главных глаз и центральным телом.
Однако большинство волокон, выходящих из них, связано с передними ассоциативными органами — грибовидными телами. Эти последние состоят из глобулюсов, гломерулюсов и поперечной комиссуры, так называемого мостика. Гломерулюсы являются вторичными зрительными центрами, в которых перекрещиваются многочисленные нервные волокна, выходящие из первичных зрительных ганглиев. Комиссура грибовидных тел, или мостик, таким образом, тесно связана с вторичными зрительными ганглиями побочных глаз и представляет, следовательно, вторичную и переднюю зрительную комиссуру.
Тритоцеребральная часть мозга, слагающаяся из хелицеральных ганглиев, лежит по бокам пищевода. От протоцеребрума она отделяется соединительнотканной перегородкой, пронизанной кровеносными сосудами. Ее комиссуры — передняя и задняя — проходят под пищеводом, т. е. посторально.
Таково в общих чертах строение мозга Lycosldae и других бродячих пауков. У форм, живущих на тенетах, имеются, однако, значительные упрощения. Так, у Araneidae и Linyphiidae зрительные ганглии верхнего зрительного пути сильно упрощены. Значительные отличия имеются и в строении нижнего зрительного пути.
Грибовидные тела развиты очень слабо; их мостик представлен сравнительно слабой передней зрительной комиссурой. У большинства тенетных форм (Agelenidae, Theridiidae, Gnaphosidae, Dysderidae, Amaurobiidae и мн. др.) зрительные центры развиты еще слабее, а грибовидные тела отсутствуют совсем.

Подглоточная нервная масса
Большая подглоточная нервная масса (см. рис. 70, 72) состоит из двух неравных частей: передней, очень широкой и звездообразной, слагающейся из ганглиев головогруди, и задней, очень узкой и вытянутой, образованной ганглиями брюшка. Передняя часть имеет пять парных боковых лопастей (см. рис. 70). От них начинаются мощные нервы конечностей. Таким образом, эта часть подглоточной массы имеет отчетливую наружную метамерию, и ее сегментарный состав совершенно ясен. В нее входят ганглии педипальпального и четырех следующих сегментов.
Общее число невромеров подглоточной массы различно у низших и высших пауков.
У Liphistius их 17, как у Amblypygi, и, следовательно, на долю брюшка (опистосомы) приходится 12 невромеров. Эти последние иннервируют стебелек (VII сегмент) и 11 сегментов брюшка. Замечательно, что все 17 парных ганглиев подглоточной массы Liphistius разделены еще ясными промежутками.
Все прочие пауки (топ-19 пауков для домашнего разведения) имеют в подглоточной массе 12 невромеров. Таким образом, у них сохраняется только 7 опистосомальных невромеров, что связано, конечно, со значительной редукцией задних сегментов брюшка. Кроме того, и концентрация у них гораздо сильнее; все ганглии плотно соединяются в несегментированную внешне массу и разделены лишь тонкими соединительнотканными прослойками, в которых проходят межузелковые артерии (arteriae interganglionares) и иногда трахеи
По данным Шимкевича (1887) и Морина (1888), у зародышей высших пауков сначала закладывается гораздо больше брюшных ганглиев, но затем часть из них исчезает. Так, у эмбриона Theridium сначала ясно различимы 16 парных ганглиев брюшной цепочки, из коих лишь 12 передних затем образуют подглоточную нервную массу (см. рис. 108, А).
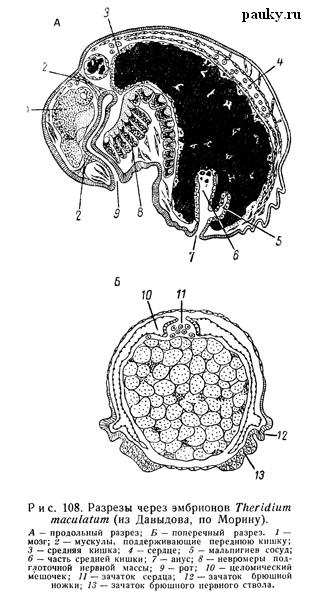
У мигаломорфного паука Chilobrachys вскоре после вылупления из яйца, помимо 12 невромеров головогруди, найдено в брюшке еще 5 парных, исчезающих затем ганглиев, т. е. в брюшной нервной цепочке на этой стадии насчитывается 17 невромеров, сходно со взрослыми Liphistiidae (Buxton, 1917).
Периферическая нервная система
Периферическая нервная система у всех пауков, по-видимому, более или менее сходна. От верхнего переднего края мозга берут начало толстые зрительные нервы (n. optici — см. рис. 70, 72), основания которых сближены в медиальной плоскости. Нервы хелицер (n. chelicerae — см. рис. 72, А, Б) отходят под ними от нижнего тритоцеребрального отдела мозга и проходят над пищеводом. Рядом с ними начинаются и парные нервы симпатической системы.
От пяти парных боковых лопастей передней части подглоточной нервной массы отходят мощные нервы конечностей головогруди (см. рис. 70). Нервные волокна, берущие начало от невромеров заднего суженного участка подглоточной массы, соединяются сзади в мощный непарный нервный ствол. Он проходит сквозь стебелек в брюшко, где сразу же делится на два ствола.

Но еще раньше от него ответвляется пара небольших нервов VII сегмента (стебелька). Каждый из парных стволов в брюшке последовательно дает три ветви. Первые две ветви принадлежат VIII сегменту. Из них одна тянется вдоль медиальной стороны легкого (первого легкого у Mygalomorphae), а другая — в косом направлении над легким к латеральному эндостерниту VIII сегмента. Третья ветвь принадлежит IX сегменту и проходит назад вдоль брюшной стенки тела.
Главные нервные стволы достигают медиальных эндостернитов IX сегмента, где разделяются каждый на три ветви: нерв передних паутинных бородавок и крибеллума (X сегмент), нерв задних и внутренних бородавок (XI сегмент) и нерв XII сегмента, направляющийся к нижней стенке клоаки и к области анального отверстия.
У Liphistiomorphae, судя по полному развитию у них задних сегментов брюшка и судя по числу невромеров, входящих в состав подглоточной нервной массы, должно быть еще несколько пар сегментарных нервов.
Симпатическая нервная система
У пауков описан лишь церебральный отдел так называемой симпатической нервной системы, связанный с задним отделом мозга (тритоцеребрум) и представленный непарной и парной частями.
Непарная часть состоит из стомодеального мостика и отходящего от него рострального нерва. Мостик, описанный Гольмгреном (1910), представляет собой небольшой ганглий и лежит в мозге непосредственно над пищеводом, охватывая его сверху в виде подковообразной перемычки. Последнее обстоятельство, так же как тесная связь элементов мостика с невропилем хелицеральных ганглиев и ганглиозной корой мозга, сначала дало повод считать его преоральной мозговой комиссурой.
Однако, как выяснилось позднее (Holmgren, 1920), он гомологичен паре стоматогастрических нервов и непарному фронтальному ганглию многоножек и насекомых. Своей комиссурообразной формой мостик обязан резкому укорочению входящих в его состав стоматогастрических нервов и включению ганглия в состав мозга. Интересно, что у эмбрионов Trochosa стомодеальнын мостик лежит еще свободно поверх мозга, как это свойственно, например, Limulus и многим Myriapoda, и лишь позднее включается в мозг (Holmgren, 1920).
От передней стороны мостика берет начало непарный, так называемый ростральный нерв (n. rostralis — см. рис. 72, 73). Проходя поверх пищевода, он тянется вперед к мускулам глотки. Назад от мостика тянется непарный возвратный нерв (n. recurrens), который несколько сзади от мозга соединяется с маленьким супраинтестинальным ганглием.
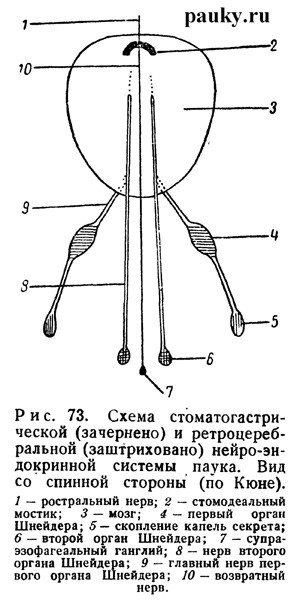
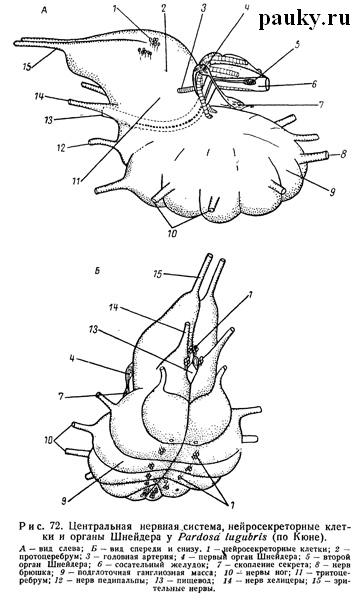
Парная часть «стоматогастрической системы» частично была известна еще в конце прошлого столетия (Schneider, 1892). Она представлена двумя парами маленьких нервов — главными и глоточными (или фарингеальными) нервами. Обе пары начинаются от задней поверхности тритоцеребрума и тянутся назад и вверх возле пищевода (рис. 72). Сзади они соединяются соответственно с двумя парами небольших железистых органов, которые еще недавно обозначались как «стоматогастрические ганглии».
Нейросекреция
В центральном нервном аппарате пауков имеются нейросекреторные клетки двух типов, открытые Лежандром (Legendre, 1954), а также Габе (Gabe, 1954). Они не имеют нейритов или аксонов в отличие от нейросекреторных клеток позвоночных. Среди Chelicerata раньше они были известны лишь у Limulus (Scharrer, 1941). Сейчас морфология системы нейросекреторных клеток пауков хорошо известна благодаря прекрасному исследованию Кюне (Kuhne, 1959).
У трех изученных им родов — Tegenaria, Trochosa и Araneus нейросекреторная система оказалась весьма сходной. В мозге нейросекреторные элементы «типа А» лежат парными группами (по 1 — 10 клеток) в периферическом слое ганглиозных клеток (рис. 72, А). В области протоцеребрума имеются две парные группы, в тритоцеребруме тоже две; по одной паре имеют ганглии педипальп, ходных ног и опистосомальная нервная масса. Таким образом, нейросекреторные клетки располагаются в общем метамерно.

Нейросекреторные клетки «типа А», крупные и округлые, обнаруживают специфическое отношение к некоторым красителям и содержат характерный секрет. Более мелкие, звездчатые нейросекреторные клетки «типа В» окружают повсюду нейропиль мозга и содержат зернистый секрет, накапливающийся между нейропилем и слоем ганглиозных клеток и нередко наблюдающийся между аксонами нервных волокон.
Другая часть нейросекреторных клеток сосредоточена в «стоматогастрических ганглиях», которые состоят преимущественно из них, т. е. представляют собой инкреторные железистые органы. Ввиду этого теперь они обозначаются нейтральным названием органов Шнейдера (Legendre, 1956; Kuhne, 1959). Те, которые иннервируются главными нервами, были открыты еще Шнейдером.
Это передние органы Шнейдера. Они охватывают в виде колечка верхние концы внутренних выростов эндостернита и состоят из нейросекреторных и мелких нервных клеток. От каждого органа назад тянется продолжение нерва, связанное со своего рода резервуаром с каплеобразным секретом, который отсюда изливается в гемолимфу в области коксальных желез (см. рис. 72, 73). Задними органами Шнейдера называются инкреторные органы, иннервируемые глоточными нервами (рис. 72, 73). Из них секрет поступает непосредственно в кровь (Kuhne, 1959).
В нервах, связанных с органами Шнейдера, можно видеть капельки нейросекрета, хотя и не в таком большом количестве, как у насекомых в межцеребральной части протоцеребрума и кардиальных телах. Поэтому Кюне полагает, что у пауков нейросекрет поступает как в нервную систему, так и в гемолимфу.
В настоящее время еще не удается обнаружить существенное сходство в морфологии «стоматогастрических нервов» у пауков, скорпионов, сольпуг и жгутоногих. Подмечено, что в период наступления у животных половой зрелости нейросекреторные клетки, в общем богаты секретом. Возможно, их деятельность стимулирует созревание половых продуктов и половую активность животного. Возможно также, что нейросекреция имеет отношение к процессам линьки.





